Как называются женские половые гормоны выделяемые фолликулярными клетками яичника
Фолликулярные клетки яичника женщины выделяют гормоны
Гормоны яичников. Эстрогены
В настоящее время установлено. что яичники выделяют следующие гормоны: 1) эстрогены; 2) прогестерон; 3) релаксин. Кроме того, выделяются и андрогены симпатикотропными клетками, описанными Бергером. Изолировано 3 эстрогенных гормона. эстрадиол, эстрон, эстриол. Все эти эстрогены имеют характерное ядро—холестран с тремя двойными связями в первом кольце. Самым активным гормоном является а-эстрадиол, затем следуют эстрон и эстриол. а-Эстрадиол в 10 раз активнее эстрона и в 50 раз — эстриола. Из мочи беременной зайчихи были изолированы 5 стероидов эстрогенных: аквилин и его изомер гиппулин, эквилинин, b-эстрадиол и b-17-дегидроэквилин.
Эстрадиол (фолликулярный гормон, дегидрофолликулин, дегидроэстрол) появляется в организме в форме двух изомеров: а-эстрадиол и b-эстрадиол. Натуральный и активный гормон циркулирует в крови в форме комплекса с b-липопротеинами (связь происходит на уровне печени, будучи в зависимости от функционального состояния последней) названными эстропротеинами; этот комплекс используется на уровне периферийных тканей. Образование эстропротеинов находится в зависимости и от уровни кортизола крови, с которым эстрогены связаны активной частью белковой молекулы.
Эстриол был изолирован в 1930 г, Doisy и Marian из мочи беременной женщины. Долгое время эстриол считали неактивным метаболитом. Исследования показали, что этот гормон активный. Эстриол появляется в яичниках, яичках, плаценте и т. д. Выделение его с мочой растет во время беременности и уменьшается во время кормления, в период климакса или после кастрации. Введение АКТГ повышает выделение эстрогенов у кастрированных. В период беременности в наибольшем количестве выделяется эстриол. Если выделение эстриола в этот период ниже 1000 мкг, то это может быть обусловлено дисфункцией плаценты и смертью плода. При токсикозе беременности выделение эстриола понижается, в то время как секреция гопадотропинов остается повышенной. В яичниках при этом происходят следующие процессы: появляется эстрадиол, превращающийся в эстрон, а затем в эстриол. В ткани яичника радиоактивный тестостерон превращается в радиоактивный эстрон, эстрадиол и эстриол.
Исследования показывают. что эстриол является главным эстрогеном, циркулирующим в плазме человека; он соединяется с глюкуроновой кислотой. Кроме того, 2/3 этого гормона связаны с липопротеинами (эстропротеины). Эстриол, как и другие эстрогены, вызывает утолщение и ороговение клеток влагалища. Он вызывает расширение и открытие отверстия влагалища. Восстановление отверстия влагалища служит тестом для дозирования эстриола (1 единица эстриола = 0,16 мкг у-эстрона). Он стимулирует рост матки, по в меньшей мере, чем эстрадиол и эстрон (соотношение активности этих гормонов: эстрон—100, эстриол—300, эстрадиол—40). Рост матки происходит за счет гидратации ее тканей. В то время как эстрон и эстрадиол стимулируют развитие мышц и эндометрия матки, эстриол действует особенно на уровне стромы матки и производит расширение шейного канала.
Нарастание эстриола перед родами вызывает расслабление и расширение шейки матки и влагалища. У женщин с ригидностью матки и дисменореей содержание эстриола понижено. Выделение эстрогенов имеет место и в коре надпочечников, в плаценте и в яичках.
Freed и Soskin считают, что эстрогены не действуют на эндометрий. Гранулезная зона яйцеклетки выделяет эстрон. Интерстициальные клетки яичника также выделяют эстрогены или содержат предшественники последнего. Количество циркулируемого эстрогена трудно определить, так как 75% этого гормона не извлекается эфиром, ибо находится в соединении с протеинами.
Общее количество эстрогенов. выделенное во время менструального цикла, равно 1,3—1,6 мг%. Среднее ежедневное выделение эстрогенов у здоровых женщин варьирует между 15 и 100 мкг. У мужчин их выделение меньше. Во время беременности выделяется большое количество эстрогенов (Риск).
Эстрогены коры надпочечников — эстрон и аллопрегненолон — синтезируются в ретикулярной зоне. Эти эстрогены вызывают феминизацию. У больных с опухолью надпочечников были изолированы большие количества эстрадиола, эстрона, эстриола, прогестерона и эквилина, вызывающие гинекомастию.
Автор - Татьяна Петрова.
Яичник функционирует циклично и, следовательно, его строение зависит от фазы менструального/овариального цикла или наличия беременности; поэтому, здесь сначала будет изложена общая структурная организация яичника и описаны все его структурные компоненты, а затем - изменения, происходящие в процессе овариального цикла и беременности
Яичник условно разделяется на корковое и мозговое вещество.- кроковое вещество содержит почти все компоненты паренхимы, между которыми располагаются прослойки рыхлой соединительной ткани (строма)
- мозговое вещество образовано рыхлой соединительной тканью (строма), а элементов паренхимы в нем практически нет, имеются только интерстициальные клетки
СТРОМАплотная строма - белочная оболочка покрывает яичник снаружи, образована из плотной волокнистой неоформленной соединительной ткани; на поверхности белочной оболочки имеется однослойный кубический эпителий
мягкая строма - интерстициальная рыхлая влолкнистая соединительная ткань
ПАРЕНХИМА образована:- фолликулами яичника на разных уровнях развития
- желтыми телами на разных уровнях развития
- интерстициальными клетками
- белыми телами - проросшими соединительной тканью остатками фолликулов и желтых тел
- атретическими (подвергающимися обратному развитию - атрезии) фолликулами
интерстициальные клетки располагаются диффузно в строме, с преобладанием во внутренней теке и мозговом веществе; продуцируют предшественников эстрогеновых гормонов, этими предшественниками являются андрогены
фолликулы яичника образованы ооцитом I порядка, окруженным фолликулярными клетками; в зависимости от степени зрелости различают следующие виды фолликулов:- примордиальные фолликулы - самые незрелые; имеются в большом количестве; располагаются, в основном, в поверхностных отделах; состоят из ооцита I порядка, окруженного одним слоем плоских фолликулярных клеток,в процессе роста превращается в
- первичный фолликул. ктороый только что вступил в рост; состоит из ооцита I порядка, который окружен одним или несколькими слоями кубических фолликулярных клетокпревращается во
- вторичный фолликул - это более зрелая форма, по сравенению с первичным фолликулом; состоит из ооцита I порядка, окруженного несолькими слоями фолликулярных клеток, между которыми образуются полости, заполненные фолликулярной жидкостью;в процессе дальнейшего роста превращается в
- третичный фолликул имеет в центре полость, заполненную жидкостью, а ооцит I порядка и фолликулярные клетки расолагаются у стенки фолликула, фолликулярные клетки со всех сторон покрывают ооцит I порядка, имеется сформированная тека; в дальнейшем превращается в
- зрелый фолликул - это готовый к овуляции фолликул; принципиально имеет такое же строение, что и третичный фолликул, только большего размера
- атретический фолликул - фолликул, подвергающийся обратному развитию или атрезии, в таком фолликуле имеются признаки гибели ооцита (сморщенная блестящая оболочка, уменьшенный объем цитоплазмы и кариопикноз), происходит гибель фолликулярных клеток и уменьшение объема фолликулярной жидкости; атретический фолликул со временем прорастает соединительной тканью (белое тело)
фолликулярные клетки находятся внутри фолликулов, вырабатывают эстрогеновые гормоны (эстрадиол, эстрон, эстриол) из предшественников, которые синтезируются интерстициальными клетками вне фолликула
тека - наружная соединительнотканная оболочка фолликула формируется в процессе роста фолликула и подразделяется на:- наружную теку. представляющую собой рыхлую соединительную ткань с небольшим количеством сосудов
- внутреннюю теку. состоящую из рыхлой соединительной ткани, большого количества интерстициальных клеток и множества сосудов
- образование предшественников эстрогенов - осуществляется интерстициальными клетками вне фолликула, затем эти предшественники поступают в фолликул
- образование самих эстрогенов осуществляется фолликулярными клетками внутри фолликула, куда поступают предшественники эстрогенов и из них фолликулярные клетки синтезируют эстрогеновые гормоны
ОВУЛЯЦИЯ - это разрыв стенки фолликула и выход него ооцита I порядка; овуляция индуцируется очень большой концентрацией лютеинизирующего гормона, который стимулирует синтез и высвобождение протеолитических ферментов фибробластами, макрофагами, тучными клетками, нейтрофилами, имеющимися в соединительнотканной оболочке фолликула, что способствует истончению и разрыву стенки фолликула; в процессе овуляции из фолликула выходит ооцит и вытекает фолликулярная жидкость, а затем на месте фолликула из фолликулярных клеток образуется желтое тело
желтое тело образуется на месте овулировавшего фолликула из фолликулярных клеток, которые в процессе образования желтого тела превращаются в лютеоциты
желтое тело состоит из лютеолцитов, которые не образуют каких-либо специфических структур, и прослоек рыхлой соединительной тканью, богато кровоснабжается; лютеоциты вырабатывают гормон желтого тела - прогестерон
если не происходит имплантации бластоцисты, то желтое тело функционирует с 14 до 25 день менструального цикла и быстро подвергается обратному развитию (с 25 по 28 день); если произошла имплантация, то под действием хорионического гонадотропина, вырабатываемого трофобластом, происходит еще большее развитие желтого тела (желтое тело беременности) и оно функционирует всю беременность, и окончательно атрофируется только после прекращения лактации
в развитии и функционировании желтого тела выделяют несколько стадий:- стадия васкуляризации и пролиферации - происходит прорастание кровеносных сосудов внутрь овулировавшего фолликула, делятся фолликулярные клетки
- стадия железистого метаморфоза - фолликулярные клетки превращаются в лютеоциты
- стадия расцвета - активное функционирование желтого тела
- стадия инволюции - обратное развитие, сопровождающееся апоптозом лютеоцитов и разрастанием соединительной ткани
белое тело - это проросшие соединительной тканью остатки от фолликулов и желтых тел
ЯИЧНИК ПРИ БЕРЕМЕННОСТИ не осуществляет циклических изменений и не содержит растущих фолликулов, имеются примордиальные фолликулы, белые тела и хорошо развитое (до 5 см в диаметре) желтое тело беременности; в другом яичнике - только примордиальные фолликулы и белые тела
Яичники
Яичники — это парный орган, расположенный по бокам от матки. рядом с ампулярными отделами маточных труб, размеры их колеблются от 1,5 до 5 см. Яичники выполняют две важнейших функции — хранение и ежемесячное созревание половых клеток, а также эндокринную функцию: они выделяют в кровоток женские половые гормоны, основными из которых являются эстрогены и прогестерон, а также небольшое количество андрогенов (мужских половых гормонов). Две эти функции делают яичники центральным звеном репродуктивной системы женщины. В них хранится информация о будущем потомстве, в результате чего женщина может иметь генетически собственных детей, и именно они делают женщину женщиной как внешне, так и психологически, обеспечивая так называемый гонадный и фенотипический пол. Аналогичным органом у мужчин являются яички. Органом — регулятором яичников является гипофиз, который находится в головном мозге и синтезирует фолликуло-стимулирующий (ФСГ) и лютеинизирующий гормоны (ЛГ). В свою очередь, гланым органом-мишенью для гормонов яичников является матка. Другие органы мишени: молочные железы, влагалище, мочевой пузырь. кожа, волосяные фолликулы, кости, сосуды и многие другие. Поэтому недостаток половых гормонов проявляется очень многообразно (см. Климактерический синдром ).
Яичники имеют два слоя — корковое вещество и строму. В корковом веществе расположены множественные различных размеров «пузырьки» (фолликулы), содержащие яйцеклетки (женские половые клетки) на разных стадиях созревания. Размеры фолликулов колеблются от микроскопических до 1,5מ см. Последние называются предовуляторными. Как ясно из их названия, они являются наиболее зрелыми, готовыми к овуляции — выходу готовой к оплодотворению зрелой яйцеклетки. Обычно в каждом цикле у женщины созревает один такой фолликул, гораздо реже — два, и крайне редко — три. Однако, в программах ЭКО (экстракорпорального оплодотворения), где применяются стимуляторы овуляции, таких фолликулов может созреть чуть ли не 20. Фолликул достигает своей зрелости примерно к 14 дню (+/- 5 дней) 28-дневного менструального цикла. В этот момент происходит резкий выброс большого количества гормонов гипофиза и яичников, в результате чего стенка фолликула разрывается и яйцеклетка выходит в брюшную полость (происходит овуляция). В период своего роста (до момента овуляции) фолликул становится эндокринной железой — он вырабатывает большое количество женских половых гормонов — преимущественно, эстрогены. После овуляции фолликул резко меняет свою структуру и трансформируется в т. н. желтое тело — оно начинает вырабатывать прогестерон — гормон второй фазы менструального цикла. Прогестерон — это и основной гормон беременности. Поэтому если происходит оплодотворение овулировавшей яйцеклетки, то желтое тело увеличивается в размерах и поддерживает уровень прогестерона до тех пор, пока эту функцию не возьмет на себя плацента (т. е. до 12 недель беременности). Если же оплодотворение не произошло, желтое тело постепенно угасает, и к 28 дню количество гормонов резко снижается — начинаются очередные месячные. Далее цикл повторяется сначала — и так на протяжении всего репродуктивного периода в жизни женщины.
На протяжении жизни женщины яичник подвергается возрастным изменениям, как никакой другой орган. Количество половых клеток в яичнике зародыша женского пола на 10 неделе внутриутробного периода развития составляет около миллиона. Это их максимальное число. На протяжении всей остальной жизни яйцеклетки постепенно гибнут, и к 45 годам их уже нет ни одной. Репродуктивный (детородный) период у женщин короче, чем у мужчин, и длится в среднем от 15 до 45 лет. В этот период яйцеклетки циклически созревают, гормоны усиленно вырабатываются, и возможна беременность. Принципиально важным является то, что новых яйцеклеток у женщин (в отличие от сперматозоидов мужчин) не появляется, а все время расходуются только уже имеющиеся. Основным выводом является то, что репродуктивное здоровье женщины начинает формироваться «в утробе матери», и беречь его надо всю жизнь, поскольку все неблагоприятные воздействия яичник «запоминает», что может отразиться на способности к зачатию и на качестве потомства. Как говорится, «берегите честь смолоду».
Ниже приведен список заболеваний, которые могут поражать яичник:
- Недостаточность функции яичников (гипо-, нормо- и гипергонадотропная), принимающая самые разнообразные клинические формы, но проявляющаяся прежде всего ановуляцией на фоне нарушения гормональной функции
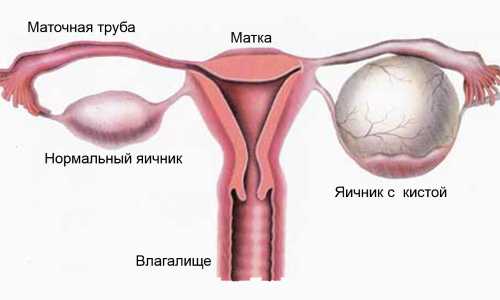
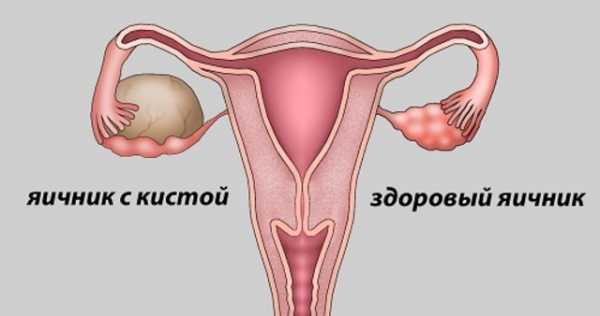
- Опухолевидные образования яичников (т. н. функциональные, фолликулярные, ретенционные кисты)
- Доброкачественные опухоли яичников (т. н. «кисты»)
- Эндометриоз яичников
- Рак яичников, лимфома.
Андрос - урология, гинекология,эндокринология
Источники: http://medicalplanet.su/gormonalnie_narushenia/296.html, http://www.histol.ru/tables/ovary.htm, http://www.andros.ru/ganat/org_ovarium.html
Комментариев пока нет!kalendarberemennost.ru
ПОИСК
Фолликулостимулирующий гормон — гормон передней доли гипофиза, вызывающий рост фолликулов яичника и секрецию эстрогенов. [c.468]Эстрогенные или женские половые гормоны выделяются яичниками. В последних секретируются два типа гормонов фолликулярные, нли эстрогенные, вырабатываемые клетками созревающего фолликула и прогесте-нальный гормон (прогестерон), секретируемый клетками желтого тела, образующегося на месте разорвавшегося фолликула. Эстрогенные гормоны обеспечивают развитие женских половых органов, вызывают периодическое наступление овуляции, разрастание слизистой оболочки матки, подготовку ее к воспринятию яйца и развитию плода. Эстрогенные гормоны вызывают также развитие вторичных женских половых признаков. У животных эстрогенные гормоны вызывают течку. Течка вызывается также введением кастрированным животным (мышам) экстрактов яичника, и на этом явлении основан важнейший метод испытания активности женских половых гормонов, разработанный Алленом и Дойзи (1923 г.). У мышей и крыс отдельные стадии полового цикла распознаются при рассматривании под микроскопом мазка, взятого из влагалища, содержащего ороговевшие эпителиальные клетки. По методике авторов, исследуемое вещество растворяют в масле и вводят подкожно тремя порциями в течение 24 ч. Одной мышиной единицей (МЕ) называют наименьшее количество вещества, которое у 70% мышей вызывает явления эструса. По международному соглашению 1939 г. для стандартизации женских гормонов используется кристаллический препарат, под названием эстрон, 0,17 которого представляет собой 1 международную единицу (1 МЕ) эстрогенной активности. Чувствительность метода позволяет определить 0,1 у эстрона чувствительность повышается при интрава-гинальном введении. [c.587]
По мере развития первичного фолликула клетки гранулезы секретируют жидкость, содержащую эстроген — один из женских половых гормонов. В фолликуле обрузуется пространство, заполненное жидкостью, — антрум. Теперь фолликул называют вторичным фолликулом. Эстроген стимулирует рост фолликула, который в конечном счете превращается в зрелый фолликул, известный под названием граафова нузьфь-ка. Этот пузырек достигает 1 см в диаметре. Он содержит ооцит второго порядка и первое полярное тельце, образующееся при первом мейо-тическом делении ооцита первого порядка (рис. 21.35). Ооцит второго порядка содержит гаплоидное число хромосом. Второе мейотическое деление достигает стадии метафазы, но не [c.81]Женские половые гормоны эстрадиол и эстриол вырабатываются клетками созревающего фолликула. Рост и созревание фолликула регулируются фолликулостимулирующим гормоном гипофиза (ФСГ). Под влиянием лютеинизирующе-го гормона гипофиза (ИКСГ — интерстициальные клетки, стимулирующий гормон) усиливается секреция эстрогенов. [c.95]
Физиол. роль ФСГ заключается в регуляции ф-ции половых желез. При этом он действует совместно с лютеинизирующим гормоном. ФСГ стимулирует рост и созревание фолликулов в яичниках у самок, развитие и созревание сперматозоидов в семенниках у самцов, усиливает секрецию половыми железами эстрогенов и авдрюгенов, повышает чувствительность половых желез к лютеинизирующему гормону, [c.113]
Физиол. роль ХГ состоит в поддержании норма 1ЬНого развития беременности. ХГ стимулирует продуцирование желтым телом, образующимся в яичниках при беременности, стероидных гормонов прогестерона и эстрогенов, что является необходимым условием для сохранения беременности. Кроме того, ХГ участвует в регуляции др. процессов, связанных с нормальным протеканием беременности, в т. ч. препятствует отторжению зародыша в материнском организме, влияя на иммунные процессы. При введении ХГ в организм человека или животных наблюдается эффект, аналогичный действию лютеинизирующего гормона. У небеременных женщин ХГ вызывает овуляцию в созревших фолликулах и образование желтых тел в яичниках. У мужчии ХГ стимулирует секрецию тестостерона половыми железами. [c.307]
Прогестерон — это основной гормон желтого тела, эндокринной железы, развивающейся в яичнике из фолликула после высвобождения яйцеклетки (овуляции). Прогестерон образуется также в надпочечниках, семенниках и плаценте. Он подвергается чрезвычайно быстрым превращениям в организме, в основном путем восстановления в соответствующий спирт, который затем образует конъюгированные соединения и выводится в виде глюкуронидов [уравнение (12-12)]. Восстановление двойной связи в кольце А прогестерона приводит к полной потере биологической активности, что указывает на важную роль а,р-нена-сыщенного кетона в проявлении гормонального действия. Прогестерон функционирует как гормон, необходимый для поддержания беременности, и, кроме того, наряду с эстрогенами он участвует в регуляции менструального цикла. [c.584]
Чем индущ1руется первоначальный рост отдельных примордиальных фолликулов, неизвестно, ио полагают, что гормональная стимуляция не имеет к этому отношения. С другой стороны, продолжение развития таких фолликулов, вероятно, зависит от гонадотропных гормонов гипофиза [в основном от фолликулостимулирующего гормона (ФСГ)] и от эстрогенов, секретируемых [c.33]
Надо сказать, что пока практически можно оказывать влияние только на женщин. Как известно, примерно четырехнедельный цикл деятельности яичника заключается в том, что в нем развивается фолликул-пузырек величиной с горошину, в котором содержится яйцеклетка. Из приведенной выше схемы следует, что и прогестерон (гормон желтого тела) и эстрадиол (женский половой гормон) тормозят образование в гипофизе гормонов ФСГ и ГСРСТ необходимых для развития яйцеклетки. Если в крови искусственно повысить концентрацию эстрогенов (женских половых гормонов) или гестагенов (гормонов желтого тела), то можно полностью прервать менструальный цикл. Без ФСГ не может созреть фолликул, а без ГСРСТ не произой- [c.346]
Прогестерон выделяется в яичнике желтым телом (корпус лутеум), органом, образованным клетками, покрывающими фолликулу после разрыва и выталкивания яичка. Регулирование менструального цикла осуществляется эстрогенами совместно с прогестероном. У женщин менструальный цикл состоит из периодической подготовки матки к беременности, осуществляемой в две фазы. Первая фаза протекает под гормональным влиянием эстрогенов и заключается в росте или пролиферации (делении клеток) слизистой оболочки матки. На протяжении второй, секреторной, фазы гормон желтого тела подготавливает слизистую оболочку матки к имплантации оплодотворенного яичка. В том случае, если оплодотворение не имело места, слизистая оболочка в значительной степени дегенерирует и возвращается в непролиферированное состояние. [c.297]
Физиологические взаимоотношения между гормонами гипофиза и гормонами яичника весьма сложны, ибо гормоны яичника, в свою очередь, до некоторой степени регулируют секреторную активность гипофиза . У овариэктомированных животных выделяется большее количество гормона, стимулирующего рост фолликул, чем у нормальных животных при введении эстрогенов выделение гормона понижается. Более того, выделение лутеинизирующего гормона регулируется прогестероном. Чувствительное равновесие стимулирующих и тормозящих эффектов [c.298]
Фоллитропин и лютропин образуют группу гонадотропных гормонов. Фоллитропин регулирует созревание фолликулов у женщин и сперматогенез у мужчин лютропин стимулирует секрецию эстрогенов и прогестерона, разрыв фолликулов и образование желтого тела у женщин, развитие интерстициальной ткани и секрецию тестостерона у мужчин. Оба гормона являются сложными белками — гликопротеинами состоят из двух субъединиц — аир (каждая из них в отдельности лишена биологической активности). Специфичность действия гормонов зависит от р-субъединиц, которые имеют разное строение, а а-субъединицы — сходное. [c.383]
Клетки гранулезы развивающегося фолликула начинают вырабатьшать женский половой гормон эстроген. Эстроген представляет собой стероидный гормон, продукция которого в первой половине цикла постоянно увеличивается в соответствии с ростом фолликула (рис. 21.44). Эстроген воздействует на два органа-мишени — матку и переднюю долю гипофиза. В матке он стимулирует восстановление и развитие ее выстилки — эндометрия, что необходимо для подготовки матки к возможной беременности, поскольку в этом случае зародьш должен имплантироваться в эндометрий. В передней доле гипофиза эстроген подавляет секрецию ФСГ по принципу отрицательной обратной связи, тем самым препятствуя развитию других фолликулов таким [c.82]
Лютеинизирующий гормон стимулирует созревание фолликулов, синтез эстрогенных гормонов, желтого тела (после разрыва фолликула) и прогестерона. В семенниках усиливает инкрецию тестостерона. [c.65]
Стероидные гормоны отличаются между собой по количеству атомов углерода. К стероидам с 21 атомом относятся гормоны коры надпочечников и гормон желтого тела (прогестерон) группа с 19 атомами углерода представлена мужским половым гормоном стероиды с 18 атомами углерода являются эстрогенными гормонами (например, фолликулярные гормоны, которые приводят к созреванию фолликулов и стимуляции овоцитогене-за и циклические изменения слизистой оболочки матки и влагалища). [c.205]
У млекопитающих синтез прогестерона в яичниках контролируется гипофизом. По сигналу от гипоталамуса гипофиз секретирует фолли-кулостимулирующий гормон (ФСГ, один из гонадотропных гормонов, который стимулирует созревание фолликула). Примерно в середине менструального цикла резко возрастает секреция другого гипофизарного гонадотропина — лутеинизирующего гормона (ЛГ) одновременно снижается секреция ФСГ. Под влиянием ЛГ фолликул разрывается, яйцо высвобождается и образуется желтое тело. Главная функция желтого тела — синтез прогестерона. Под влиянием прогестерона и эстрогенов, также образующихся в яичниках (гл. XII), матка подготавливается для имплантации оплодотворенного яйца. Если оплодотворения не происходит, то желтое тело и вызываемые прогестероном изменения подвергаются инволюции, а часть слизистой матки слущивается во время менструашш. [c.69]
Ооциты первого порядка у новорожденной девочки останавливаются в профазе 1 мейоза (подобно ооцитам первого порядка у амфибий), и большая их часть окружена одним слоем фолликулярных клеток такой ооцит вместе с этими фолликулярными клетками представляет собой примордиальный фолликул (рис. 15-28) За некоторое время до рождения небольшая доля примордиальных фолликулов периодически начинает расти, превращаясь в развивающиеся фолликулы их клетки увеличиваются и размножаются, образуя вокруг ооцита первого порядка многослойную оболочку сам ооцит, оставаясь в профазе 1, растет, и у него формируются zona pellu ida и кортикальные гранулы (рис. 15-30). В некоторых из развивающихся фолликулов образуется наполненная жидкостью полость, или antrum, при этом они превращаются в антральные фолликулы (рис. 15-31). Чем индуцируется первоначальный рост отдельных примордиальных фолликулов и превращение их в развивающиеся фолликулы, пока неясно. Однако продолжение развития, вероятно, зависит от гонадотропных гормонов гипофиза [в основном от фолликулостимулирующего гормона (ФСГ)] и от эстрогенов, секретируемых самими фолликулярными клетками. [c.35]
В детстве концентрация половых гормонов низка, хотя экзогенные гонадотропины увеличивают их продукцию. Следовательно, незрелые яичники обладают способностью синтезировать эстроген. Существует предположение, что у неполовозрелых девочек имеющиеся у них в небольшом количестве половые стероиды подавляют образование гонадотропинов, а в пубертатном возрасте гипоталамогипо-физарная система становится менее чувствительной к ингибирующему действию этих гормонов. В период полового созревания начинается импульсная секреция ГнРГ (гонадолиберина) под влиянием ЛГ резко повышается уровень образования яичниковых гормонов, а под влиянием ФСГ, главного стимулятора секреции эстрогенов, происходит созревание фолликулов и наступает овуляция. [c.238]
chem21.info
Женские половые гормоны
Женские половые гормоны можно разделить на две группы – собственно половые гормоны, вырабатываемые в яичниках, надпочечниках и гормоны гипоталамуса, влияющие на выработку гормональных веществ в половых органах, надпочечниках.
Женские половые гормоны гипоталамуса
Эти вещества называются гонадотропинами – от слов «гонадо» — половые органы и «тропин» — влияющий. Именно они регулируют выработку собственно женских половых гормонов и гормональные циклы.
Типы секреции гонадотропинов
В нормальном овуляторном цикле выделяют фолликулярную и лютеиновую фазы выделения женских половых гормонов. Фолликулярная фаза начинается с момента менструации и достигает кульминации в момент предовуляторного пика повышения концентрации лютропина. Лютеиновая фаза начинается с предовуляторного повышения содержания лютропина и заканчивается в первый день менструации.
Снижение женских половых гормонов эстрадиола и прогестерона вследствие регресса желтого тела предыдущего цикла по механизму обратной связи инициирует повышение содержания ФСГ,которое стимулирует рост фолликула, секрецию эстрадиола. Основной признак — рост фолликула и секрецию эстрадиола — объясняют двугонадотропной (лютропин и ФСГ) двуклеточной (тека-клетки и гранулезные клетки) теорией развития фолликулов в яичнике. В соответствии с ней клетки фолликулов яичника обладают некоторыми функциями, в результате которых под действием лютропина тека-клетки синтезируют андрогены (андростендион, тестостерон), а затем под влиянием ФСГ в гранулезных клетках происходит превращение этих андрогенов в эстрогены (андростендиона — в эстрон, тестостерона — в эстрадиол). Первоначально, при более низких концентрациях эстрадиола, существует эффект влияния обратной связи на готовую к выделению форму лютропина из пула гонадотропинов в гонадотрофах гипофиза. Поскольку содержание женского полового гормона эстрадиола позднее (в фолликулярной фазе) повышается, существует положительная обратная связь, обеспечивающая выделение гонадотропинов из запасов, что приводит к резкому повышению концентрации лютропина и овуляции. Последняя происходит в период от 36 до 44 ч после начала повышения содержания лютропина в середине цикла. При применении фармакологических доз гестагенов, содержащихся в противозачаточных таблетках, отмечают глубокий эффект влияния обратной связи на гонадотропин-рилизинг-гормон (ГнРГ), поэтому из пула гонадотропинов не выделяется ни один гормон, таким образом, овуляция блокируется.
Во время лютеиновой фазы повышенные концентрации циркулирующего эстрадиола и прогестерона по механизму обратной связи существенно подавляют выработку лютропина и ФСГ. Это угнетение сохраняется почти до окончания лютеиновой фазы, пока не снизится содержание прогестерона и эстрадиола в результате регрессии желтого тела и не наступит беременность. Совокупный эффект состоит в слабом повышении сывороточной концентрации ФСГ, которое инициирует рост новых фолликулов для следующего цикла. Продолжительность функциональной инволюции желтого тела такова, что при отсутствии беременности менструация обычно наступает через 14 дней после достижения пикового содержания лютропина.
Гипоталамус
Гипоталамус выделяет различных малых пептидов или биогенных аминов, влияющих на репродуктивный цикл. Все они оказывают специфическое действие на секрецию гормонов передней доли гипофиза. К ним относят ГнРГ, тиреотропин-рилизинг-гормон (ТРГ), соматотропин-ингибирующий фактор или соматостатин, кортикотропин-рилизинг фактор, пролактин-ингибирующий фактор (ПИФ). В этой статье о женских половых гормонах рассмотрены лишь ГнРГ и ПИФ.
ГнРГ — декапептид, который, в первую очередь, синтезируется в дугообразных ядрах. Он отвечает за синтез и выделение женских половых гормонов — лютропина и ФСГ. Поскольку он в большей степени стимулирует выделение ЛГ, чем ФСГ, его иногда называют лютропин-рилизинг-гормоном или лютропин-рилизинг фактором (ЛРФ). Вероятно, ФСГ и лютропин присутствуют в гонадотрофах гипофиза в двух различных формах — выделяемой и запасной. ГнРГ проникает в гипофиз через его портальные сосуды и стимулирует синтез ФСГ и лютропина, хранящегося в гонадотрофах. ГнРГ последовательно активирует и трансформирует эти молекулы в выделяемые формы. Кроме того, он может вызывать немедленное выделение лютропина и ФСГ в кровоток. Во время недавнего исследования были обнаружены рецепторы к ГнРГ в других тканях (включая яичник), что свидетельствует, что ГнРГ может оказывать прямое влияние на функционирование яичника и повышение женских половых гормонов.
На протяжении менструального цикла ГнРГ секретируется в пульсовом режиме. Частота его выделения, косвенно определенная по пульсовым выделениям лютропина, варьирует примерно от одного импульса на каждые 90 мин в ранней фолликулярной фазе до одного импульса на каждые 60-70 мин непосредственно в предовуляторном периоде. Во время лютеиновой фазы частота импульсов падает, а их амплитуда повышается. Установлены значительные вариации у отдельных лиц.
Внутривенное и подкожное введение экзогенного пульсового ГнРГ применяют для индукции овуляции у женщин, у которых она отсутствует в результате дисфункции гипоталамуса. Длительная (непульсовая) инфузия ГнРГ приводит к обратимому угнетению секреции гонадотропинов через процесс дезактивации или десенсибилизации гонадотрофов гипофиза. Это отражает основной механизм действия агонистов ГнРГ (нонапептидов, содержащих только девять аминокислот), которые успешно применяют для лечения заболеваний, связанных с нарушением гормональной регуляции яичников (эндометриоза, лейомиомы, гирсутизма, преждевременного полового созревания).
Секрецию ГнРГ контролируют несколько механизмов. Эстрадиол, вероятно, усиливает выделение гипоталамусом ГнРГ и способствует индукции выброса лютропина в середине цикла путем повышения выработки ГнРГ или усиления ответа гипофиза на декапептид. Гонадотропины оказывают угнетающее действие на выделение ГнРГ. Кроме того, одну из основных регулирующих ролей играют катехоламины. Дофамин, синтезируемый в дугообразных и паравентрикулярных ядрах, может оказывать прямое угнетающее действие на секрецию ГнРГ через бугорково-воронковый тракт, входящий в срединное возвышение. Серотонин, вероятно, также угнетает пульсовое выделение ГнРГ, а норадреналин стимулирует его. Эндогенные опиоиды подавляют секрецию ГнРГ гипоталамусом с помощью пути, который частично регулируется стероидными гормонами яичников.
Гипоталамус синтезирует ПИФ, осуществляющий постоянное угнетение выделения женского полового гормона пролактина из лактотрофов. Некоторые фармакологические средства (например, хлорпромазин), воздействующие на дофаминергические механизмы, влияют на выделение пролактина. Собственно дофамин, секретируемый в портальные сосуды гипофиза нейронами гипоталамуса, угнетает выделение пролактина непосредственно в аденогипофизе. На основании наблюдений предполагают, что дофамин гипоталамуса может быть основным ПИФ. В дополнение к регуляции выделения пролактина ПИФ гипоталамус может синтезировать пролактин-рилизинг факторы, вызывающие мощный и быстрый выброс пролактина при определенных условиях (например, при стимуляции молочных желез во время кормления грудью). До 2008 г. биохимическая характеристика ПИФ и пролактин-рилизинг факторов отсутствовала. ТРГ также способствует стимуляции выделения пролактина. Этим феноменом можно объяснить связь между первичным гипотиреозом, сопровождающимся вторичным повышением концентрации ТРГ, и гиперпролактинемией. Установлено, что белок-предшественник ГнРГ — ГнРГ-ассоциированный пептид — сильный ингибитор секреции пролактина и усилитель выделения гонадотропина. Это свидетельствует, что ГнРГ-ассоциированный пептид также может быть физиологическим ПИФ, и объясняет обратную связь между секрецией гонадотропина и пролактина, обнаруживаемую при многих состояниях
Эстрогены
Эстрогены – основные женские половые гормоны. В ранней фолликулярной фазе концентрация циркулирующего эстрадиола относительно низкая. Примерно за неделю до овуляции его содержание начинает повышаться, сначала медленно, затем быстро. Превращение тестостерона в эстрадиол в гранулезной клетке фолликула происходит посредством ароматизации. Содержание эстрогенов достигает максимального значения за день до пикового повышения концентрации лютропина в середине цикла, после чего вплоть до овуляции происходит его выраженное и резкое снижение. В лютеиновой фазе содержание эстрадиола достигает максимума на 5-7-й день после овуляции, уровень женского полового гормона возвращается к исходному значению незадолго до менструации. Секреция эстрона яичником значительно ниже, чем секреция эстрадиола, но она происходит схожим образом. Эстрон в основном образуется из андростендиона под действием ароматазы.
Прогестагены
Во время развития фолликула яичник секретирует прогестерон и 17a-гидроксипрогестерон в очень малых количествах. Основной объем женского полового гормона прогестерона образуется на периферии при превращении прегненолона и прегненолона сульфата надпочечников. Незадолго до овуляции неразорвавшийся, но лютеинизированный граафов фолликул начинает синтезировать повышенное количество прогестерона. Примерно в это же время отмечают повышение содержания сывороточного 17a-гидроксипрогестерона. Повышение базальной температуры по времени связано с центральным действием прогестерона. Как и в ситуации с эстрадиолом, секреция прогестагенов желтым телом достигает максимума на 5-7-й день после овуляции, возвращается к исходной незадолго до менструации. Если должна наступить беременность, содержание прогестерона и, соответственно, базальная температура остаются повышенными.
Андрогены
Несмотря на то, что это мужские гормональные вещества, в организме женщины они выполняют роль женских половых гормонов — и яичники, и надпочечники секретируют малые количества тестостерона, но его большая часть образуется при метаболизме андростендиона, который также секретируется и яичниками, и надпочечниками. Примерно в середине цикла отмечают повышение концентрации андростендиона в крови, что отражает его повышенную секрецию фолликулом. В лютеиновой фазе регистрируют второе повышение содержания андростендиона, которое отражает его повышенную секрецию желтым телом. Надпочечники, как и кортизол, секретируют андростендион в суточном ритме. Яичник вырабатывает малые количества дигидротестостерона (ДГТ), но его основной объем образуется при превращении андростендиона, тестостерона. Большую часть дегидроэпиандростерона (ДГЭА) и фактически весь ДГЭА-сульфат (ДГЭАС), которые являются слабыми андрогенами, секретируют надпочечники, хотя малые количества ДГЭА секретирует яичник.
Связывающие белки сыворотки
Циркулирующие эстрогены и андрогены соединяются преимущественно со специфическими глобулинами, связывающими половые гормоны, или сывороточным альбумином. Оставшаяся фракция половых гормонов несвязанная (свободная), и именно она биологически активна. Неясно, доступны ли связанные с сывороточными белками (например, с альбумином) половые гормоны для тканевого захвата и утилизации. Синтез глобулинов, связывающих половые гормоны, в печени усиливают эстрогены, гормоны щитовидной железы, а тестостерон угнетает его.
Пролактин
Женский половой гормон пролактин секретируется лактотрофами. В отличие от других пептидных гормонов, синтезируемых аденогипофизом, гипоталамус оказывает подавляющее действие на выделение гипофизом пролактина. Период полураспада циркулирующего пролактина составляет около 20-30 мин. В дополнение к лактогенному эффекту на протяжении овуляторного цикла пролактин может влиять на функции гипоталамуса, гипофиза и яичников (особенно при хронической гиперпролактинемии).
Концентрация пролактина в сыворотке во время нормального менструального цикла изменяется незначительно. Как сывороточное содержание пролактина, так и его секреция в ответ на действие ТРГ в лютеиновой фазе повышается немного больше, чем в середине фолликулярной фазы цикла. Повышение концентрации циркулирующего эстрадиола и прогестерона способствует усилению выработки пролактина. Выделение этого женского полового гормона изменяется на протяжении дня, наибольшую концентрацию регистрируют во время сна.
Пролактин участвует в контроле стероидогенеза в яичниках. Его концентрация в фолликулярной жидкости значительно изменяется во время роста фолликула. Самое высокое содержание пролактина обнаруживают в мелких фолликулах в ранней фолликулярной фазе. Концентрация гормона в фолликулярной жидкости обратно пропорциональна продукции прогестерона. Кроме того, гиперпролактинемия может привести к нарушению секреции гонадотропинов. Несмотря на это, физиологическая роль пролактина, как женского полового гормона, во время нормального менструального цикла до настоящего времени четко не установлена.
Полезно:
surgeryzone.net
ПОИСК
В женских половых железах — яичниках — вырабатываются гормоны в созревающих фолликулах и желтом теле. Как по физиологическому действию, так и по химическому строению гормоны фолликула и желтого тела различаются между собой. Для нормального развития женского организма необходимы оба эти гормона. [c.372] Наиболее активным из этих соединений оказался эстрадиол, который и рассматривается как первичный гормон фолликулов яичника. [c.373]Гормоны фолликула (эстрогены) [c.453]
Эстрадиол — гормон, вырабатываемый клетками фолликул в яичниках, содержащих созревшие яйца. Интерстициальные клетки семенников вырабатывают один из мужских половых гормонов — тестостерон. Он превращается в андростерон. Эти соединения имеют формулы [c.420]
Основной андроген, или мужской половой гормон, — это тестостерон, образующийся из прегненолона путем отщепления боковой цепи при С-17. Образование тестостерона составляет у мужчин 6—10 мг/сут, у женщин также синтезируется тестостерон, но в небольших количествах ( 0,4 мг). Любопытно, что тестостерон служит предшественником женских половых гормонов. В крови тестостерон циркулирует в комплексе с р-глобулином и оказывает воздействие на ряд органов-мишеней, в том числе и на органы воспроизведения. Наиболее заметный эффект тестостерона — это стимуляция роста волос на лице. Вместе с тем тестостерон вызывает преждевременную гибель волосяных фолликул на голове у генетически предрасположенных индивидуумов. В результате у лысых мужчин обычно растет прекрасная борода, причем волосяные фолликулы бороды при пересадке на темя остаются устойчивыми [c.589]
Фолликулостимулирующий гормон (ФСГ) сти мулирует созревание фолликулов и сперматогенез. [c.149]
Гормоны фолликула. Жид кость, находящаяся в фолликуле, содеря ит по меньшей мере два гормона — эстрон и эстрадиол. Эстрон (теелин) был первым гормоном, выделенным из фолликулярной жидкости, однако эстрадиол (дигидротеелин) оказался более активным соединением, и, возможно, именно он является основным [c.357]
Эстрогенные или женские половые гормоны выделяются яичниками. В последних секретируются два типа гормонов фолликулярные, нли эстрогенные, вырабатываемые клетками созревающего фолликула и прогесте-нальный гормон (прогестерон), секретируемый клетками желтого тела, образующегося на месте разорвавшегося фолликула. Эстрогенные гормоны обеспечивают развитие женских половых органов, вызывают периодическое наступление овуляции, разрастание слизистой оболочки матки, подготовку ее к воспринятию яйца и развитию плода. Эстрогенные гормоны вызывают также развитие вторичных женских половых признаков. У животных эстрогенные гормоны вызывают течку. Течка вызывается также введением кастрированным животным (мышам) экстрактов яичника, и на этом явлении основан важнейший метод испытания активности женских половых гормонов, разработанный Алленом и Дойзи (1923 г.). У мышей и крыс отдельные стадии полового цикла распознаются при рассматривании под микроскопом мазка, взятого из влагалища, содержащего ороговевшие эпителиальные клетки. По методике авторов, исследуемое вещество растворяют в масле и вводят подкожно тремя порциями в течение 24 ч. Одной мышиной единицей (МЕ) называют наименьшее количество вещества, которое у 70% мышей вызывает явления эструса. По международному соглашению 1939 г. для стандартизации женских гормонов используется кристаллический препарат, под названием эстрон, 0,17 которого представляет собой 1 международную единицу (1 МЕ) эстрогенной активности. Чувствительность метода позволяет определить 0,1 у эстрона чувствительность повышается при интрава-гинальном введении. [c.587]
Биосинтез Л г осуществпяется из а- и -субъединиц, к рые образуются раздельно из соответствующих высокомол белков-предшественников, аминокислотная последовательность к-рых кодируется разл генами Образование зрелых полипептидных цепей происходит в результате специфич протеолитич расщепления белков-предшественников У женских особей Л г стимулирует овуляцию, разрыв фолликула с освобождением яйцеклетки и формирование желтого тела а также секрецию прогестерона В мужском организме Л г стимулирует ф-цию интерстициальных клеток семенников, в т ч секрецию ими мужского полового гормона тестостерона Снижение секреции Л г приводит к нарушению детородной ф-ции и бесплодию Л г выделяют из гипофизов животных и человека [c.619]
Физиол. роль ФСГ заключается в регуляции ф-ции половых желез. При этом он действует совместно с лютеинизирующим гормоном. ФСГ стимулирует рост и созревание фолликулов в яичниках у самок, развитие и созревание сперматозоидов в семенниках у самцов, усиливает секрецию половыми железами эстрогенов и авдрюгенов, повышает чувствительность половых желез к лютеинизирующему гормону, [c.113]
Физиол. роль ХГ состоит в поддержании норма 1ЬНого развития беременности. ХГ стимулирует продуцирование желтым телом, образующимся в яичниках при беременности, стероидных гормонов прогестерона и эстрогенов, что является необходимым условием для сохранения беременности. Кроме того, ХГ участвует в регуляции др. процессов, связанных с нормальным протеканием беременности, в т. ч. препятствует отторжению зародыша в материнском организме, влияя на иммунные процессы. При введении ХГ в организм человека или животных наблюдается эффект, аналогичный действию лютеинизирующего гормона. У небеременных женщин ХГ вызывает овуляцию в созревших фолликулах и образование желтых тел в яичниках. У мужчии ХГ стимулирует секрецию тестостерона половыми железами. [c.307]
Прогестерон — это основной гормон желтого тела, эндокринной железы, развивающейся в яичнике из фолликула после высвобождения яйцеклетки (овуляции). Прогестерон образуется также в надпочечниках, семенниках и плаценте. Он подвергается чрезвычайно быстрым превращениям в организме, в основном путем восстановления в соответствующий спирт, который затем образует конъюгированные соединения и выводится в виде глюкуронидов [уравнение (12-12)]. Восстановление двойной связи в кольце А прогестерона приводит к полной потере биологической активности, что указывает на важную роль а,р-нена-сыщенного кетона в проявлении гормонального действия. Прогестерон функционирует как гормон, необходимый для поддержания беременности, и, кроме того, наряду с эстрогенами он участвует в регуляции менструального цикла. [c.584]
Щитовидная железа играет исключительно важную роль в обмене веществ. Об этом свидетельствуют резкое изменение основного обмена, наблюдаемое при нарушениях деятельности щитовидной железы, а также ряд косвенных данных, в частности обильное ее кровоснабжение несмотря на небольшую массу (20—30 г). Щитовидная железа состоит из множества особых полостей —фолликулов, заполненных вязким секретом—коллоидом. В состав коллоида входит особый йодсодержащий гликопротеин с высокой мол. массой — порядка 650000 (5000 аминокислотных остатков). Этот глико-протеин получил название йодтиреоглобулина. Он представляет собой запасную форму тироксина и трийодтиронина — основньж гормонов фолликулярной части щитовидной железы. [c.264]
Биологическое действие по сравнению с специфично. Распад происходит в результате / -захва-та, который сопровождается каскадом фотонов характеристического излучения и низкоэнергетических Оже-электронов. Значительная часть этой энергии поглощается коллоидом фолликулов, а также той частью тирео-идного эпителия, где локализованы синтез и секреция гормонов. Ядра клеток об-чучаются в меньшей степени. Вследствие этого по показателю функционального состояния железы эффективнее, чем 1, а по показателю гибели клеток он менее эффективен. [c.280]
Передняя доля гипофиза вырабатывает следующие гормоны тиреотропный гормон, стимулирующий секреторную функцию щитовидной железы адренокортикотропный гормон (АКТГ), стимулирующий функцию коры надпочечных желез лутеотропный гормон, или лактоген (пролактин), вызывающий секрецию молока в молочной железе два гонадотропных гормона (гормон, стимулирующий развитие фолликулов в яичнике, и гормон лутеинизации, регулирующий образование каротиноидов в яичнике и выработку мужских половых гормонов), а также соматотроп-ный гормон или гормон роста, контролирующий размеры тела животных. [c.447]
Чем индущ1руется первоначальный рост отдельных примордиальных фолликулов, неизвестно, ио полагают, что гормональная стимуляция не имеет к этому отношения. С другой стороны, продолжение развития таких фолликулов, вероятно, зависит от гонадотропных гормонов гипофиза [в основном от фолликулостимулирующего гормона (ФСГ)] и от эстрогенов, секретируемых [c.33]
С наступлением половой зрелости раз в месяц волна выделяемого гипофизом лютеинизирую-щего гормона (ЛГ) побуждает один антральный фолликул к созреванию ооцит первого порядка, находящийся в этом фолликуле, завершает первое деление мейоза, образуя полярное тельце н превращаясь в ооцит второго порядка. 4. Ооцит второго порядка вместе с полярным тельцем и частью окружающих фолликулярных клеток осюбождается в тот момент, когда фолликул разрывается на поверхности яичника. Ооцит второго порядка претерпевает второе деление мейоза только в том случае, если он будет оплодотворен. [c.34]
Женские половые гормоны эстрадиол и эстриол вырабатываются клетками созревающего фолликула. Рост и созревание фолликула регулируются фолликулостимулирующим гормоном гипофиза (ФСГ). Под влиянием лютеинизирующе-го гормона гипофиза (ИКСГ — интерстициальные клетки, стимулирующий гормон) усиливается секреция эстрогенов. [c.95]
Женские половые гормоны. Основным органом, вырабатывающим женские половые гормоны, являются яичники. Однако эти гормоны образуются частично в плаценте и могут синтезироваться в надпочечниках. Они были обнаружены также в моче и семенниках самцов. Основным женским половым гормоном, который вырабатывается в яичниках, является эстрадиол. Кроме эстрадиола, гормональными свойствами обладают также эстрон (фолликулин) и эстриол, которые являются производными эстрадиола. Эстрадиол синтезируется в фолликулах яичников из холестерина или ацетатов промежуточными продуктами этого синтеза являются андростен-дион и тестостерон. При введении в женский организм тестостерона последний частично превращается в эстрадиол. [c.150]
Обычно процесс воспроизводства у млекопитающих начинается в гипофизе — небольшой железе, расположенной у основания головного мозга. Гипофиз считается очень важной железой, поскольку он выделяет гормоны, регулирующие гормональную секрецию других желез. В начале цикла воспроизводства в женском организме гипофиз выделяет гормон, называемый фолликулостимулирующим гормоном (ФСГ). Этот гормон, имеющий белковую природу, стимулирует развитие яйцевых фолликул в яичниках, причем ткань фолликула выделяет в кровь эстрадиол. Эстрадиол выполняет две важные функции останавливает дальнейшее выделение ФСГ, что необходимо для предотвращения одновременного созревания большого числа яйцеклеток, и стимулирует рост ткани матки, подготавливая ее к имплантации яйцеклетки, после того [c.164]
Из экстрактов передней доли гипофиза выделено, идентифицировано и охарактеризовано шесть различных гормонов. Все эти гормоны являются белками или полипептидами. 1. Гормон роста (ГР, соматотропин) — ускоряет рост тела. 2. Адренокортикотропный гормон (АКТГ, кортикотропин) — стимулирует кору надпочечников. 3. Лактогенный гормон (ЛТГ, лютеотропный гормон, пролактин) — стимулирует и поддерживает функцию желтого тела, выделение молока. 4. Тиреотропный гормон (ТСГ, тиреоидостимулирующий гормон)—стимулирует щитовидную железу. 5. Фолликулостимулирующий гормон (ФСГ) — стимулирует созревание фолликул и сперматогенез. 6. Интерстициальные клетки стимулирующий гормон (ИКСГ) — стимулирует интерстициальные клетки яичников и семенников, побуждает овуляцию, влияет на образование желтого тела. [c.196]
ПРОГЕСТЕРОН (лютеостероп). Гормон, вырабатываемый желтым телом яичников. Поддерживает беременность, способствует закреплению оплодотворенной яйцеклетки, препятствует развитию новых фолликул и проявлению охоты у самок. Нерастворим в воде, хорошо растворим в спирте и масле. Для лечебных целей выпускается в форме масляных растворов в стеклянных ампулах. Вводится подкожно и внутримышечно при нарушениях беременности (угрожающий аборт) и для стимулирования лактации крупным животным (лошадям, коровам) по 0,01—0,04 мг и мелким (свиньям, овцам) по 0,004—0,008 мг раз в день 4—8 дней подряд. [c.246]
ФОЛЛИКУЛИН. Женский половой гормон, вырабатываемый в фолликулах яичников самок животных. Фолликулярные гормоны способствуют появлению течи и охоты у самок. Эти гормоны в виде эфиров в больших количествах выде.тгяются в моче особей мужского пола (особенно жеребцов). В животноводческой практике для стимуляции охоты и полового влечения самкам иногда вводят в орга- [c.321]
Лютропин (лютеинизирующий гормон, ЛГ) — гормон передней доли гипофиза, который вырабатывают специализированные клетки аденогипофиза. У самок он вызывает разрыв фолликула с образованием желтого тела, у самцов стимулирует развитие интерстициальной ткани и секрецию тестостерона. ЛГ — сложный белок-глтокопротеид, имеющий мол. массу у овец 40 ООО, у свиней — 100 ООО. Его молекула состоит из дву а- и Р-субъединиц. Установлена последовательность аминокислотных остатков а-субъединицы овцы, свиньи и человека и локализация пяти дисульфидных связей в а-субъединице ЛГ овцы. Структура а-субъединицы у человека отличается от ее структуры у овцы 22 аминокислотными остатками, а у свиньи—пятью. ЛГ человека укорочена с №конца на семь остатков, а свиньи — на шесть. а-Суф.е-диница ЛГ состоит из 96 аминокислотных остатков и имеет два углеродных радикала, присоединенных по остаткам аспарагина в 56-м и 82-м положениях- [c.279]
chem21.info